



James T.Rutka(鲁特卡)教授曾任世界神经外科学院院长,30多年来深耕儿童神外研究,带领团队不断突破,从脑瘤分子分型、精准治疗、新药治疗、微创治疗,为全球神经外科疑难病患儿带来希望,他新近发表了研究《Brain–Pupil Coupling Revealed Through Deep Learning of Intracranial Recordings》(通过颅内记录的深度学习揭示脑-瞳孔耦合),以下是研究简述(下篇)。
上篇:【INC国际大咖研究成果】通过颅内记录的深度学习揭示脑-瞳孔耦合(上)
01
PART
研究结果

3.1|任务表现和瞳孔测量术概述
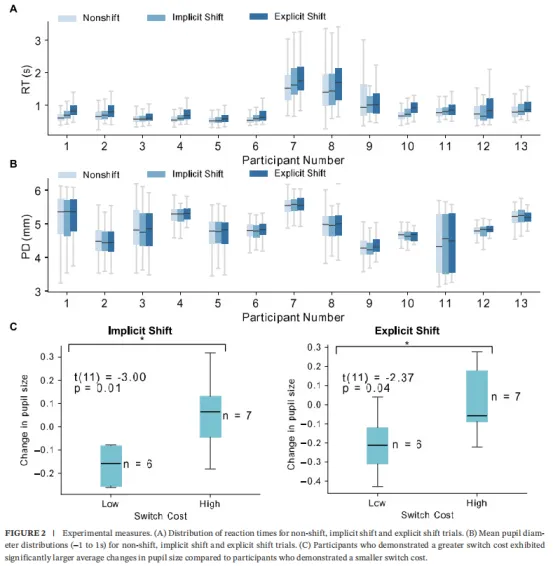
为评估任务复杂性如何影响行为表现,我们检查了不同试验类型的反应时间(图2A)。反应时间随任务复杂性增加:参与者在非转换试验中反应最快(845±536毫秒),其次是隐性转换试验(872±525毫秒),显性转换试验最慢(972±564毫秒)。LME模型确认从非转换到隐性转换试验及从非转换到显性转换试验的反应时间增加均统计显著(隐性:z=3.29,p=0.001;显性:t=12.43,p<0.001)。
为评估刺激颜色差异是否影响瞳孔反应,试验基于中位数分割(范围=0.41-0.53;图S2)二分化为“高”和“低”相对亮度组。LME模型显示相对亮度对刺激后瞳孔直径无显著影响(p=0.998,β<0.001)。
为分离任务诱发的瞳孔尺寸变化,我们比较了刺激前基线(-1至0秒)和任务诱发(0至1秒)期间跨试验类型。LME模型显示任务期间相对于基线瞳孔尺寸显著增加(z=3.002,p=0.003)。此外,任务期间瞳孔尺寸在显性转换试验中小于非转换试验(z=-2.283,p=0.022)。但非转换与隐性转换试验间瞳孔尺寸无显著差异(z=-0.526,p=0.559)。参与者间瞳孔直径分布见图2B。
为检查转换成本与瞳孔尺寸的关系,我们首先计算每位参与者的转换成本作为转换与非转换试验平均反应时间之差。参与者随后使用中位数分割分为低和高转换成本组。我们分别对隐性和显性转换条件比较这两组间的平均瞳孔尺寸变化。跨隐性和显性转换条件,t检验显示较高转换成本的参与者比较低转换成本者表现出显著更大的平均瞳孔尺寸(隐性:t(11)=-3.00,p=0.014;显性:t(11)=-2.37,p=0.037;图2C)。所有试验条件下任务准确率保持较高。儿童非转换试验平均达到98.17%(标准差:1.90%),隐性转换试验98.65%(标准差:1.54%),显性转换试验97.80%(标准差:2.42%)。LME模型显示隐性转换与非转换试验间准确率无显著差异(z=0.682,p=0.502),显性转换与非转换试验间也无显著差异(z=-0.537,p=0.596)。
3.2|瞳孔动态与任务表现相关
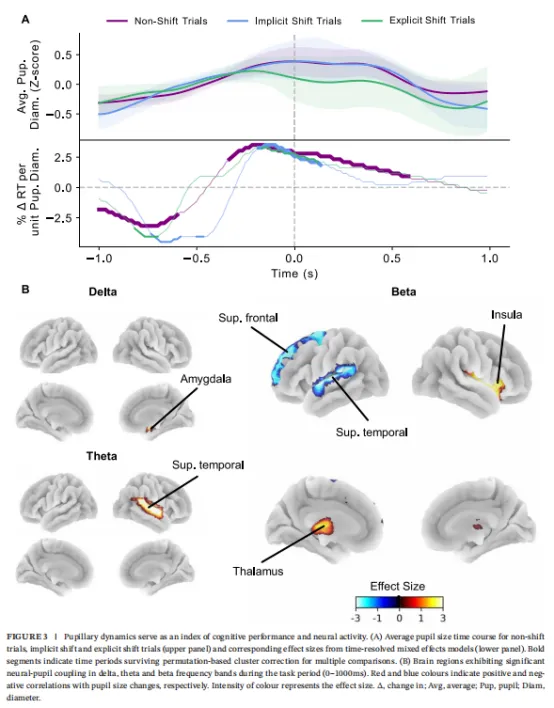
在注意定势转换任务期间,瞳孔尺寸与反应时间的关系因任务阶段和试验条件而异。在刺激间期,非转换条件下瞳孔尺寸与任务表现存在显著关联。刺激间期较大的瞳孔直径与后续试验中较快的反应时间相关。此效应存在于刺激起始相对时间-1000至-580毫秒间(β̂>-3.17,p校正<0.05;图3A,下图)。相反,此期间在隐性和显性转换试验中未观察到关系。
在刺激周围和刺激后期间,瞳孔尺寸与反应时间的关系反转,较小瞳孔直径与较快反应时间相关。此效应在非转换和隐性转换试验条件中均观察到。在非转换试验中,关联从-330至880毫秒显著,最强效应出现在约-170毫秒(β̂<3.51,p校正<0.05;图3A,下图)。类似地,在隐性转换试验中,较小瞳孔直径与较快反应时间相关,但持续时间较短,从刺激起始相对时间-170至150毫秒(β̂<3.51,p校正<0.05;图3A,下图)。
3.3|脑-瞳孔相关性汇聚于任务相关网络
我们接下来探索了注意定势转换期间瞳孔动态(即神经-瞳孔耦合)的神经相关性。FDR校正后,δ、θ和β频带出现显著相关性(图3B)。瞳孔尺寸与带限功率间的正相关在右杏仁核(δ;β̂=3.87,p校正<0.06)、右颞上回(θ;β̂=3.33,p校正<0.045)、右岛叶(β;β̂=2.99,p校正<0.037)和左丘脑(β;β̂=3.14,p校正<0.037)中观察到。在杏仁核、颞上回和丘脑中,带限功率从静息到任务增加(图S3),表明较大瞳孔直径对应较大的任务诱导神经活动增加。相反,岛叶中的β功率从静息到任务减少,提示较大瞳孔与较小的任务相关岛叶活动减少相关。
瞳孔尺寸与β功率间的负相关在左额上回(β̂=-3.04,p校正<0.037)和左颞上回(β̂=-3.49,p校正<0.025)中观察到。额上回表现出任务诱导的β功率减少,表明较大瞳孔直径与较大的β活动减少相关。反之,颞上回显示β功率从静息到任务增加,提示较大瞳孔直径对应较小的任务诱导功率增加。α或γ频带未观察到显著相关性。
3.4|SEEG信号可预测瞳孔尺寸时间过程
为揭示SEEG与瞳孔动态间的非线性、时间依赖性关系,我们部署深度学习模型基于仅SEEG记录预测瞳孔尺寸时间过程。此方法能够捕获跨特征的依赖性,如耦合的空间和谱模式。特定参与者模型在完成定势转换任务两日的13名参与者中的7名上训练。
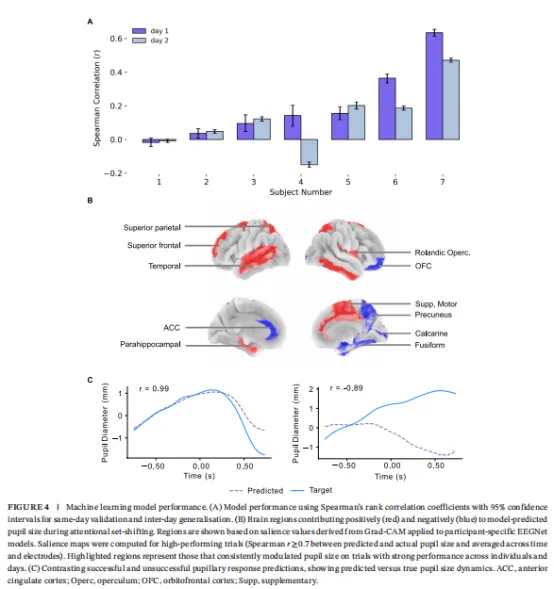
验证集和日间测试集上的模型性能显示显著的参与者特定变异性。在验证数据集上,七名模型中的六名达到统计显著结果,平均Spearman等级相关系数范围0.04至0.63(95%CI范围0.007至0.65;图4A)。为评估神经-瞳孔关系跨环境因素、睡眠模式和抗癫痫药物调整的稳定性,我们在另一天收集的数据上测试模型预测能力。日间性能显著下降,平均Spearman等级相关系数从第1日的0.24降至第2日的0.15(图4B)。
预测能力的日间稳定性跨模型变异性:基于参与者二、三和五数据训练的模型在日间保持一致性能无显著差异,而基于参与者四数据训练的模型完全丧失预测能力。两名模型表现显著下降,但保留高于机会的预测能力。
基于显著性的激活映射揭示了一个皮质和皮质下区域网络,其贡献于模型预测瞳孔尺寸(图4B)。正贡献(红色显示)在与注意控制、显著性检测、运动规划和记忆相关的区域中观察到。这些区域包括额上回、顶上小叶、辅助运动区、楔前叶、罗兰迪克岛盖、颞区、左海马旁回和右扣带。相反,负贡献(蓝色显示)在与视觉皮质(距状沟和梭状回)以及OFC、楔前叶和左ACC相关的区域中识别。成功和不良瞳孔反应预测示例如图4C所示。正Spearman等级相关系数代表成功预测,而零或负相关系数代表不良预测性能。
02
PART
讨论
瞳孔反应与认知过程耦合,为利用眼动追踪研究健康和疾病中的复杂行为提供了机会。瞳孔动态的神经基础仍不明确。通过利用认知任务期间独特颅内记录与同步瞳孔测量术,深度学习模型能够解析脑-瞳孔耦合关系。我们报告三个主要发现:(1)瞳孔尺寸与认知表现的关系随任务阶段变化,(2)特定神经网络内的任务相关活动与瞳孔波动共变,(3)在儿科癫痫患者中通过SEEG记录的神经活动可预测任务执行期间的瞳孔直径。
03
PART
意义、限制与未来方向
本研究强调了瞳孔反应作为认知处理标志物的潜力,为未来工作利用眼动追踪实时推断认知状态提供了基础。这在如癫痫儿童等人群中尤其有价值,他们常表现认知灵活性和注意力缺陷。重要的是,这些儿童经历神经活动的瞬态干扰,如来自IED(Warsi等人2023年、2022年),这些可能未被常规行为评估完全捕获。此外,识别其活动可靠预测瞳孔尺寸的特定皮质区域为理解认知表现的个体差异和可能开发针对注意力相关缺陷的靶向干预开辟了新途径。未来研究可扩展此工作,通过结合参数化变化认知需求的任务以描绘任务难度、努力和唤醒如何交互塑造瞳孔动态及其神经相关性。
本研究的一个关键限制是参与者间电极放置的变异性,这阻止了我们聚合个体间数据以训练统一模型。此变异性限制了每个模型可用的训练数据集大小,影响其性能和泛化能力。此外,我们的研究未包括所有脑和脑干区域的数据,如LC和上丘,这些是瞳孔控制网络的组成部分(Strauch等人2022年;Joshi等人2016年)。这限制了我们完全捕获瞳孔尺寸调制神经机制的能力。未来工作可利用更大SEEG数据集和更灵活架构训练组水平模型。例如,基于Transformer的模型可将空间位置编码为标记嵌入以学习跨不同电极布局参与者的可推广模式。然而,此类模型将需要substantially更大和更多样化的SEEG数据集。
总之,本研究强调了神经活动与瞳孔反应间复杂动态的关系,为认知灵活性的神经基础提供了新见解。从SEEG预测瞳孔时间过程的能力证明了深度学习在解码脑-瞳孔交互中的效用。未来研究应扩展此工作至更大、更多样化人群,使用非侵入性记录技术,如脑电图(EEG)或脑磁图(MEG),以实现更广泛适用性和泛化能力。该领域的未来研究有望开发创新方法以监测儿科癫痫的认知状态。
04
PART
关于作者

